
Le projet ISATIS
Dernière mise à jour en juillet 2018
Lancé à l'automne 2014 pour trois ans par le Laboratoire d'écotoxicologie des radionucléides (LECO), le projet ISATIS (Ionising Radiation and Biological Mechanisms) visait à analyser les dommages qu’une exposition chronique à de faibles doses de rayonnements ionisants induit sur les protéines chez les êtres vivants. Le projet s’est tout particulièrement intéressé aux mécanismes de carbonylation des protéines et à leur impact sur le fonctionnement des cellules, ceci en lien avec les dommages à l’ADN et leur réparation, les effets épigénétiques et l’apoptose (à travers d’autres projets menés au LECO) mais aussi avec les grandes fonctions des organismes. L’objectif final est une meilleure connaissance des mécanismes élémentaires impliqués dans ces réponses biologiques, notamment pour améliorer la radioprotection des espèces animales dont l’homme.
Contexte et objectifs
L’approche conventionnelle adoptée dans les études sur les effets biologiques des rayonnements ionisants considère l’ADN nucléaire comme la cible critique des dommages radio-induits (UNSCEAR, 2012). Ainsi, jusqu’à récemment, la majorité des travaux effectués dans ce domaine concernent les dommages à l’ADN et leur réparation. Or des études expérimentales (Daly et al., 2010; Krisko and Radman, 2010; Krisko et al., 2012) ont montré que les protéines, plus que l'ADN, seraient les cibles critiques des dommages causés par l'irradiation des cellules pour de nombreux modèles biologiques. Ces études démontrent que la survie de nombreux organismes est liée au niveau de dommages oxydatifs aux protéines, limitant la fonctionnalité de celles-ci, y compris l'efficacité des enzymes nécessaires pour réparer et répliquer l'ADN. La carbonylation des protéines est irréversible. Les protéines endommagées peuvent cependant être éliminées par la cellule tant que le processus de dégradation des protéines endommagées (complexes protéiques) est suffisamment actif (figure 1). À forte dose d’irradiation (i.e. 200-1000 Gy), dans certains cas, il existe une corrélation entre la production de protéines carbonylées et une baisse de la fécondité (e.g., chez des invertébrés, baisse du taux d'éclosion des œufs pondus et donc du succès reproducteur) voire la mortalité. À faible dose (< 0.1Gy), cependant, la carbonylation des protéines est un marqueur peu étudié.
Le projet ISATIS a visé à étudier le suivi de ces dommages aux protéines, particulièrement la carbonylation, peu étudiés dans ces domaines de dose. L’objectif était de mieux comprendre les mécanismes de toxicité radio-induite aux faibles doses de rayonnements ionisants en étudiant les mécanismes d’interaction entre les rayonnements ionisants et les biomolécules. Il s’est agi d’établir des corrélations entre les taux de protéines carbonylées, la nature des protéines ayant subi des dommages, l’activité de dégradation des protéines endommagées et in fine des effets « macroscopiques » type reproduction. Deux modèles de radiosensibilité distincte, le nématode Caenorhabditis elegans et le poisson Danio rerio, ont été exposés à des rayonnements g par irradiation externe à faibles et fortes doses, en aigu ou en chronique, pour une approche comparée. En parallèle, plusieurs paramètres ont été suivis : les dommages causés à l'ADN (cassures simple-brin et double-brin) et leur mécanisme de réparation ; les processus épigénétiques (changements transmissibles dans la fonction des gènes non-issus de changements de la séquence d'ADN) ; ainsi que la mort cellulaire (apoptose). L’approche choisie vise à être générique quel que soit le modèle biologique utilisé c’est à dire extrapolable sur des bases rationnelles à diverses espèces.
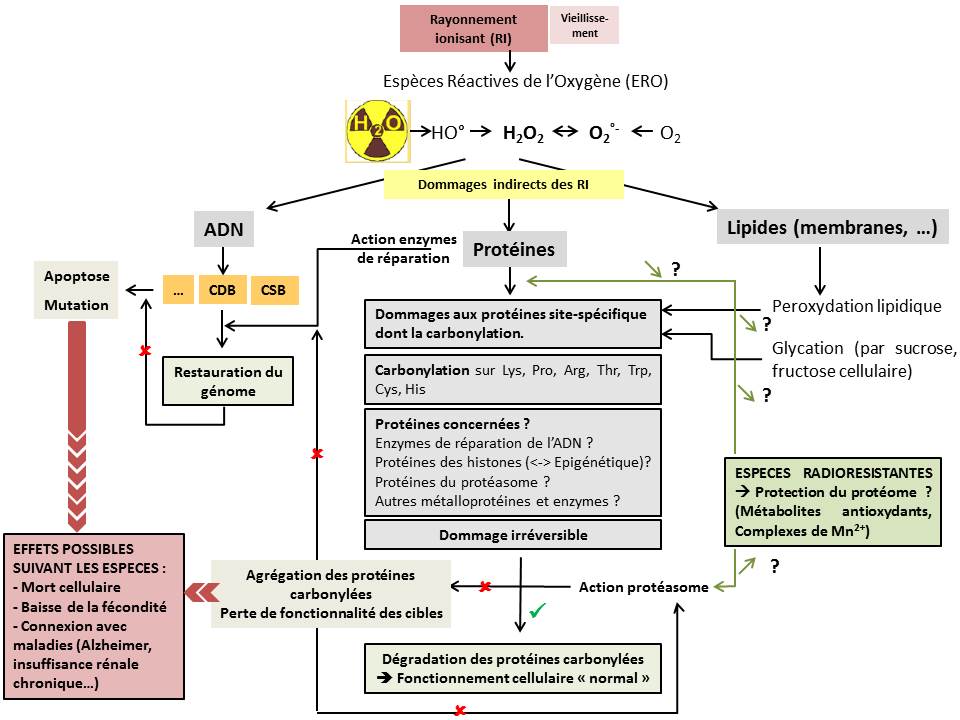
Figure 1 : Schéma de synthèse de l'implication de la carbonylation des protéines dans les processus de toxicité des rayonnements ionisants © IRSN
Déroulement du projet
Le projet ISATIS s'est appuyé sur une approche de type « biologie des systèmes » qui consistait à décrire les interactions des rayonnements ionisants avec l'ensemble des molécules-clefs dans les fonctions biologiques des organismes, notamment la reproduction : protéines, ADN, lipides... en mettant en œuvre une approche comparée entre les modèles animaux choisis.
Configuration expérimentale
Les modèles biologiques principalement utilisés au laboratoire sont des organismes modèles (C. elegans (figure 2) et D. rerio) présentant une radiosensibilité différente. Leur génome et protéome sont connus, ce qui permet d’étudier les mécanismes moléculaires et de les relier à la reproduction.
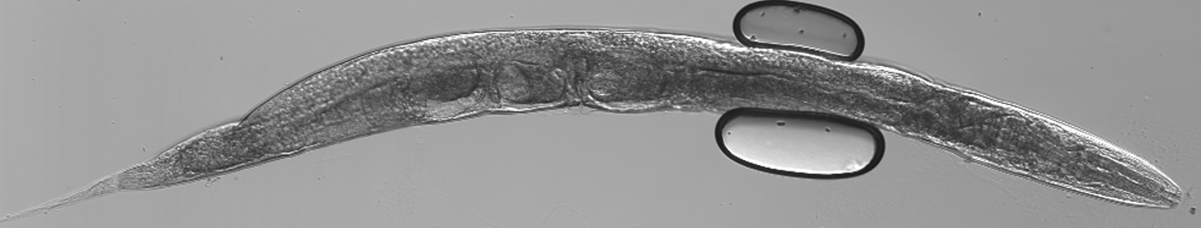
Figure 2 : Nématode C.elegans adulte © IRSN/Kuzmic M. et al.
L’exposition aux rayonnements ionisants a été réalisée par irradiation externe avec une dose maîtrisée. Ces irradiations (gamma, 137Cs) ont été effectuées dans l'irradiateur MIRRE (mini-irradiateur délivrant un débit de dose gamma externe sur des organismes de petites tailles, de quelques centaines de nGy.h-1 jusqu’à 100 mGy.h-1) puis sur la plateforme d'irradiation MICADO Lab, ainsi qu’à l’Institut Curie (irradiation aiguë).
En complément des études au laboratoire, une étude a été réalisée sur la rainette arboricole Hyla orientalis, prélevée sur différents sites de la zone d’exclusion de Tchernobyl, contaminées en 137Cs et 90Sr. Les amphibiens, tels que la rainette arboricole, représentent des organismes de choix pour plusieurs raisons : (i) ils ont une peau très perméable aux polluants, (ii) ils sont exposés aux polluants via des habitats diversifiés (aquatiques et terrestres) au cours de leur vie, (iii) ils font partie des animaux de références listés par la Commission Internationale de Protection Radiologique (CIPR) pour évaluer le risque radiologique pour l'environnement.
Analyses effectuées
- Une évaluation du taux de carbonylation des protéines et de l’activité protéolytique
Deux méthodes de mesure de la carbonylation ont été mises au point. La première méthode permet de dénombrer et d’identifier les protéines carbonylées dans l’échantillon de plusieurs individus ayant subi le même traitement. Cette méthode repose sur l’extraction et la séparation chimique des protéines, dont les carbonyles sont marqués spécifiquement. La seconde méthode permet de mesurer le niveau de carbonylation des biomolécules (protéines, ADN, ARN et lipides) dans l’organisme entier (figure 3). Elle utilise l’imagerie confocale, et un marquage multiple des biomolécules (marquage des carbonyles et marquage spécifique identifiant chaque biomolécule).
En parallèle, l’activité du protéasome (complexe protéique permettant notamment la dégradation des protéines endommagées) a été évaluée dans les différentes conditions d’irradiation pour observer une potentielle modification de la capacité cellulaire à dégrader les protéines oxydées.
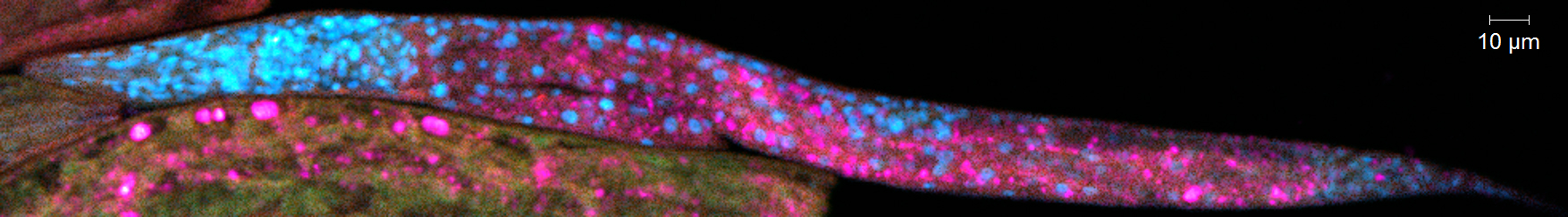
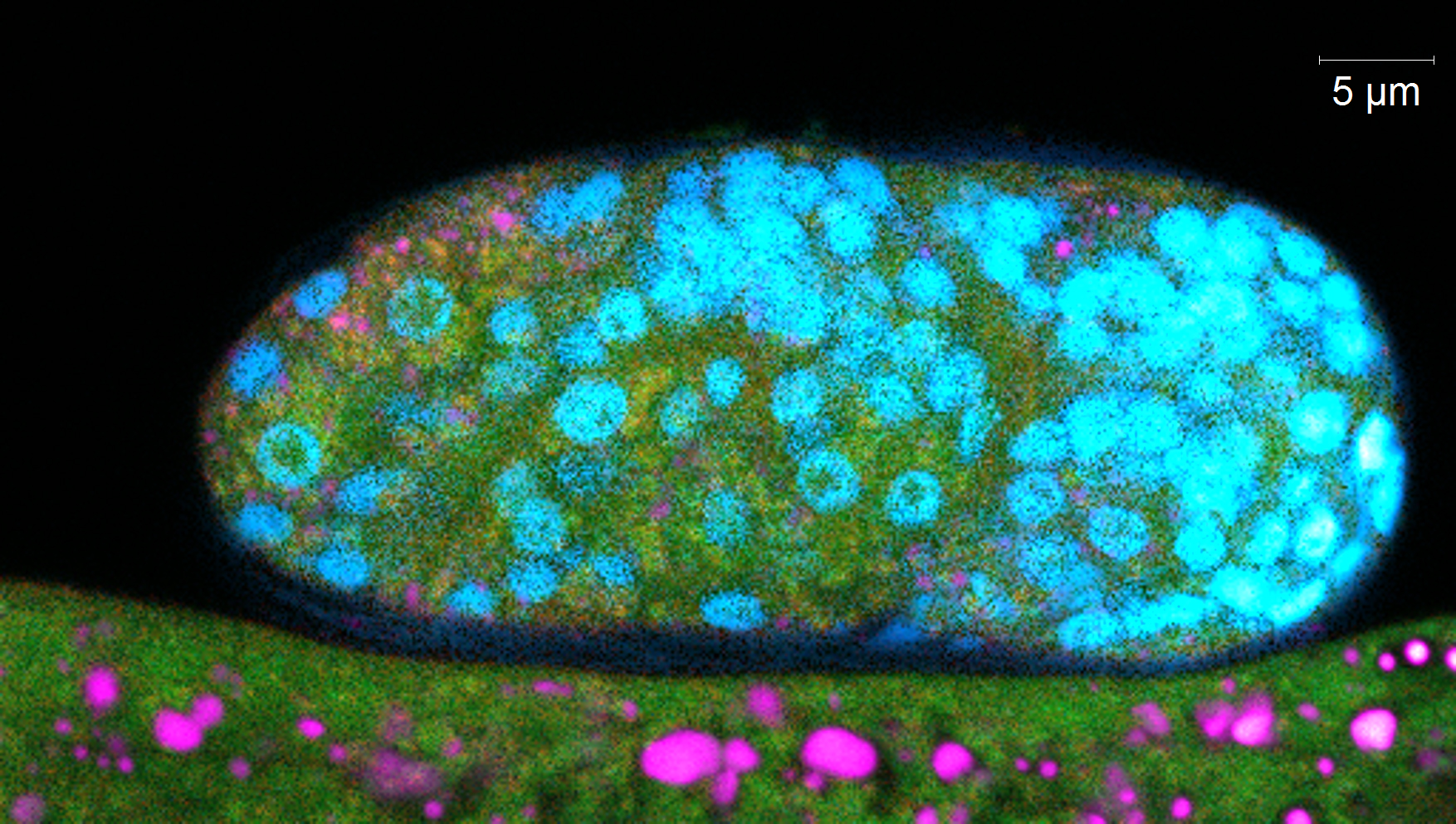
Figure 3 : Images confocales (fluorescence) de nématodes C. elegans adultes (haut) et oeufs (abs) marqués avec un marquage multiple carbonyles/lipides/ADN/protéines. © IRSN/Kuzmic M. et al.
- Une évaluation de la modulation du protéome
Une autre clé pour comprendre la radiotoxicité est d’étudier l’impact des rayonnements ionisants au niveau des protéines exprimées suite à l’irradiation. Les protéines étant impliquées dans de nombreux processus biologiques tels que l'activité protéolytique cellulaire (notamment via le protéasome), la défense antioxydante ou la réparation de l'ADN, leur étude est indispensable pour connaître les processus impliqués dans ces phénomènes de toxicité radioinduite. L’approche protéomique permet de mettre en évidence les modifications de l’expression des protéines pour un individu exposé par rapport à un individu témoin et ainsi établir un profil de modification suite à une irradiation.
- Une évaluation des dommages à l’ADN : dommages oxydatifs et réparation
La mesure des altérations de l’ADN (cassures simple-brin, cassures double-brin, bases modifiées) a été envisagée mais n’a pu être effectuée que sur D. rerio, via la méthode des comètes, au cours de projets menés au laboratoire en parallèle du projet ISATIS.
- Une évaluation des changements épigénétiques
Les changements épigénétiques ont été évalués à travers d’autres projets menés au laboratoire, à l’aide de plusieurs marqueurs, en fonction du niveau d’observation choisi et du modèle biologique, notamment la méthylation de l’ADN, qui joue un rôle dans le maintien de la structure et de la stabilité des chromosomes et dans la transcription.
- Une évaluation de l’impact sur de grandes fonctions biologiques
L’irradiation et la mesure des paramètres moléculaires et cellulaires ont été mises en regard d’effets « macroscopiques » sur les grandes fonctions biologiques : longévité et reproduction (nombre d'oeufs pondus, taux d'éclosion...).
Résultats
Sont présentés ci-après les résultats d’évaluation de la carbonylation, de l’expression des protéines et de l’activité protéolytique en réponse aux rayonnements ionisants (g, exposition 3H, sites pollués), obtenus après expositions (chronique ou aigüe) de différents organismes modèles ou environnementaux à des doses modérées. Non exhaustives, ces données sont mises en regard d’autres marqueurs moléculaires si possible mais surtout de paramètres macroscopiques tels que la reproduction ou la longévité.
(i) Lien entre la carbonylation des protéines et la reproduction après irradiation gamma aiguë ou chronique de la souche sauvage de C. elegans.
Cette étude a permis d’identifier qu’après irradiation aiguë (0-200 Gy ; 1-15 Gy.min-1), une diminution significative de la ponte totale par individu et une diminution du succès d’éclosion étaient observées à partir de 30 Gy alors qu’après irradiation chronique (0-6,5Gy ; 7-100 mGy.h-1) seule une diminution significative de la ponte totale par individu était observée mais à partir de 3,3 Gy (50 mGy.h-1 pendant 65h). De plus, il a été montré que le niveau de protéines carbonylées était augmenté après irradiation aiguë pour certaines doses (6,5 Gy notamment), mais qu’il était plutôt stable après irradiation chronique. Ce niveau de protéines carbonylées est dépendant de la quantité d’espèces réactives de l’oxygène (ERO) mais aussi de la capacité cellulaire à éliminer les protéines modifiées (activité du protéasome notamment). Suivant la nature du substrat à dégrader, 3 formes du protéasome peuvent être mises en jeu (Figure 4A). Ici la forme 20S (réputée comme affectée à la dégradation des protéines oxydées) semble avoir l’activité la plus dégradée par l’irradiation aiguë (en revanche il y a peu d’impact sur son expression) (Figure 4B). L’activité et l’expression des autres formes ATP/Ubiquitine dépendantes (30S et 26S), reflétant l’activité globale du protéasome de par leur abondance, semblent peu impactées après irradiation aiguë aux doses les plus faibles mais sont diminuées à forte dose. Après irradiation chronique au contraire, il semblerait que le protéasome, et particulièrement la forme 20S, soit stimulé.
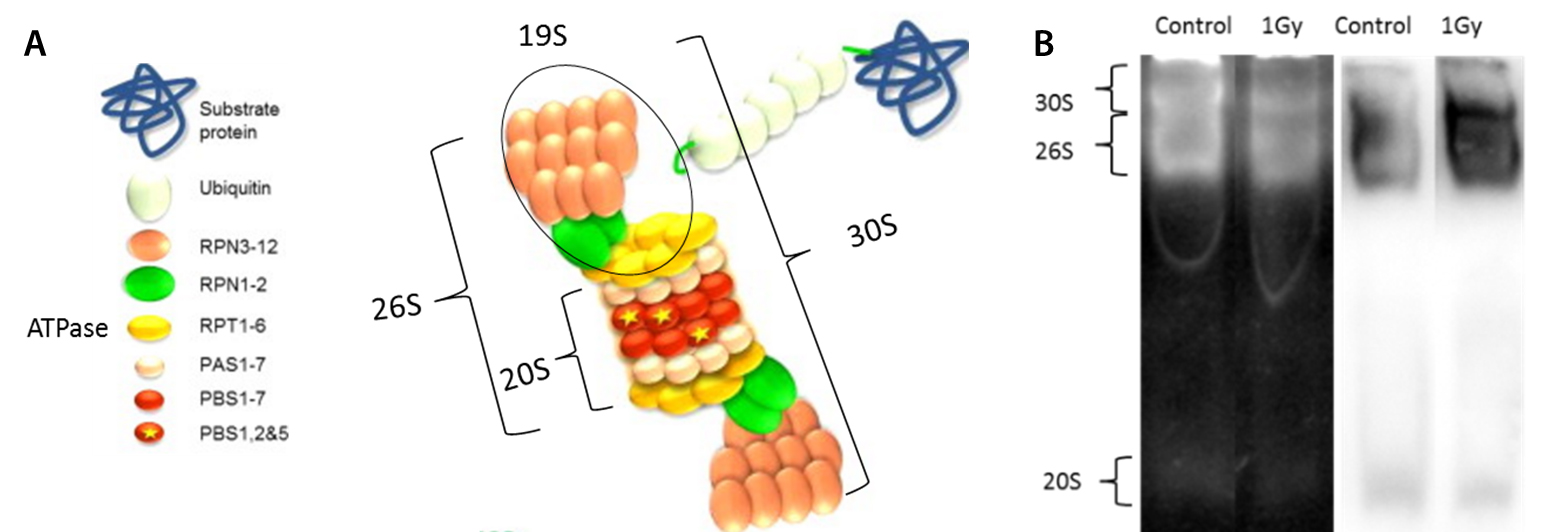
Figure 4 : A/ Schéma du protéasome et de ses sous-unités constitutives (Papaevgeniou and Chondrogianni., 2014 The ubiquitin proteasome system in C .elegans and its regulation). Unités 26S et 30S : ATP et ubiquitine dépendantes ; unité 20S : ATP et Ubiquitine indépendante. B/ Mesure sur gel natif, après séparation électrophorétique, de l’activité des protéasomes 30S, 26S et 20S (panel de gauche) et niveaux d’expression des protéasomes 30S 26S et 20S par Westernblot (panel de droite). Pour chaque ensemble d’images, à gauche : Signal issu des nématodes témoins (définis comme 100% d’activité) et à droite : signal issus des nématodes exposés à 1Gy d’irradiation aiguë (Dubois et al, Sci. Rep. 2018). © IRSN
Par ailleurs, une analyse protéomique a permis d’identifier des voies de signalisation potentiellement impliquées chez C. elegans dans la réponse aux rayonnements ionisants, en aigu et en chronique. Il a été identifié que les mécanismes sous-jacents sont distincts en aigu et en chronique. Ce résultat implique que les risques chroniques des rayonnements ionisants ne peuvent pas être simplement extrapolés à partir de données issues d’irradiation aiguë. Quelques cibles protéiques en lien avec la reproduction (ovogénèse, embryogénèse) et la réponse au stress, ainsi que l’activité du protéasome semblent être des paramètres moléculaires particulièrement intéressants à cibler lors de prochaines expérimentations, à plus faible dose ou sur plusieurs générations. En effet, ces marqueurs protéiques identifiés sont de possibles candidats pour prédire les effets de l’irradiation.
Une étude précédente avait pu mettre en évidence une influence de l’irradiation gamma externe sur le nombre de gamètes mâles (non directement mise en évidence ici à travers l’étude protéomique) ; pour approfondir les connaissances sur les mécanismes moléculaires liés à la reprotoxicité et valider les marqueurs identifiés par approche globale, il serait intéressant de réaliser une étude comparée spécifique sur la radiosensibilité des gamètes mâles et femelles chez C. elegans (morphologique et moléculaire), pour déterminer les causes du défaut de reproduction.
(ii) Lien entre la carbonylation des protéines et le vieillissement après irradiation gamma chronique du mutant glp-1 de C. elegans
Sur cette étude, nos résultats indiquent qu’une exposition chronique aux rayonnements ionisants réduit la durée de vie du mutant stérile C. elegans Glp-1 et ce quels que soient la durée d’exposition (du stade œufs à 65h ; du stade œuf à 19j), le débit de dose (7 ou 50 mGy.h-1) et la dose (0,5 ; 3,5 ou 24 Gy) testés. La question complémentaire est de savoir si les résultats seraient les mêmes avec une exposition encore plus courte (du stade œuf à 24h ou 48h), couvrant un stade de développement spécifique, par ex. diacinèse (démontré dans la littérature pour être le plus radiosensible). Au niveau de la carbonylation, elle s'est avérée être un bon biomarqueur du vieillissement puisque les nématodes âgés présentent une carbonylation plus élevée que les jeunes. L’irradiation au contraire, réduit le niveau de carbonylation quelle que soit la dose ou la durée de l’exposition. Cette diminution de la carbonylation pourrait en partie s'expliquer par une augmentation de l'activité du protéasome, déjà très actif chez ces mutants. Par ailleurs, la quantité de lipides a été étudiée et suit les mêmes tendances que la carbonylation i.e. augmentation avec l’âge, mais diminution après irradiation (Figure 5). Les lipides sont des molécules de signalisation très importantes qui constituent la membrane cellulaire ainsi qu’un stockage d'énergie. De plus, le lien entre la fonction reproductive, l'homéostasie lipidique et la longévité est décrit dans la littérature, même si les mécanismes d’impact de l’un sur l’autre ne sont pas clairement définis. Nos résultats restent donc à investiguer plus profondément pour une meilleure compréhension des connexions possibles. Quoiqu’il en soit, les niveaux de lipides et la carbonylation pourraient être des marqueurs potentiels de l'irradiation et du vieillissement, respectivement.
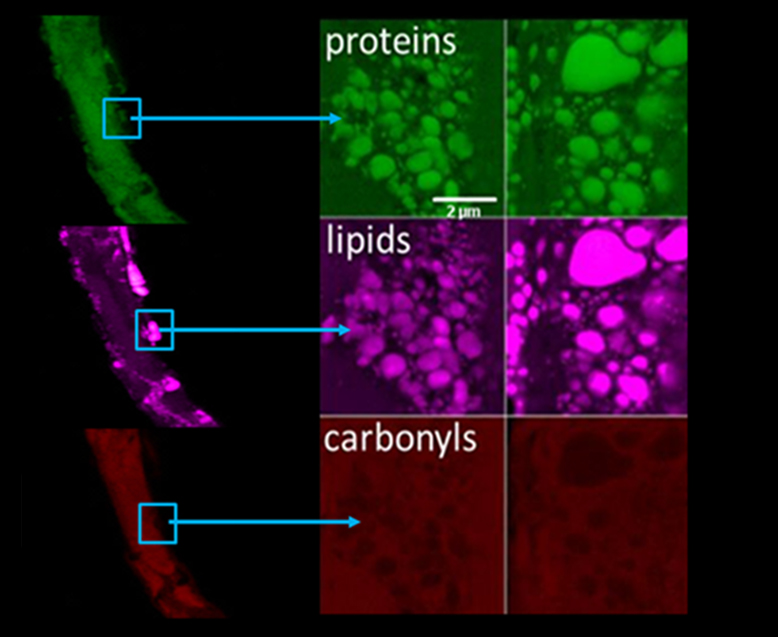
Figure 5 : Colocalisation positive entre les protéines et les lipides au stade adulte (10j), irradié 10j avec un débit de dose de 52 mGy.h-1 (dose cumulative 12,5 Gy) et au stade âgé (19j, non irradié), excluant la co-localisation avec les carbonyles (coefficient carbonyle de Pearson ~ 0,2 ) – Thèse M. Kuzmic.© IRSN
(iii) Détermination du niveau de protéines carbonylées (PC) chez Danio rerio
Après irradiation g chronique à 0,5 et 5 mGy.h-1 de juvéniles (œufs à larves), issus de parents irradiés à 0,5 et 5 mGy.h-1 dans le cadre d’une autre étude menée au laboratoire, aucun effet significatif sur le niveau de carbonylation (tendance à l’augmentation à 10j pour 0,5 et 5 mGy.h-1) n’a pu être observé après 4j et 10j d’irradiation. L’analyse de l’activité protéolytique cellulaire dans ces conditions, pourrait permettre d’apporter des éléments pour l’interprétation de cette réponse.
Après exposition à l'eau tritiée, à raison de 3,7.102 ou 3,4.103 µGy.h-1, de stades juvéniles (œuf à larves) pendant 7 et 10j pour une autre étude menée au LECO, une diminution significative du niveau de protéines carbonylées est observée à 10j par rapport aux témoins. Ceci pourrait s’expliquer par un niveau moindre d’espèces oxydatives dans le milieu (ce qui ne transparaît après analyse in vivo dans les larves de l’indice de production d’espèces réactives de l’oxygène) ou par une stimulation importante des systèmes de défense cellulaire telle la protéolyse, détruisant les protéines endommagées pour permettre leur nouvelle synthèse ; cette dernière hypothèse pourrait faire l’objet d’une étude spécifique.
(iv) Détermination du niveau de protéines carbonylées (PC) dans le muscle d’H. orientalis :
Au travers d’une autre étude menée au laboratoire, l’étude du marqueur de carbonylation et de l’expression protéique a pu être réalisée dans le muscle de grenouilles arboricoles H. orientalis issues de 3 sites dans la zone de Tchernobyl, différentiellement contaminés (en Cs et Sr) et ayant des débits de dose ambiants respectifs de 0,10 ; 3,7 et 32,4 µGy.h-1. Au niveau de la carbonylation des protéines, peu de différences sont observées mais le niveau semble diminuer chez les individus du site intermédiaire. Par ailleurs, une approche protéomique comparative a montré, sur ces mêmes grenouilles, une répression de plusieurs protéines musculaires chez les individus des deux sites les plus contaminés. La poursuite de l’analyse de ces résultats nécessite des investigations complémentaires, notamment l’identification d’un génome de référence n’existant que très partiellement pour H. orientalis pour identifier les protéines variantes et caractériser leur fonction biologique. Ils seront ensuite à mettre en regard de la dose totale (interne et externe) reçue par chaque individu.
Conclusion
Au bilan de ces analyses très diverses sur la carbonylation des protéines dans des organismes en développement irradiés au laboratoire (D. rerio, C. elegans) ou dans les tissus musculaires d’organismes prélevés dans un environnement contaminé (H. orientalis), il apparaît que ce dommage protéique, marqueur du vieillissement dans nos études, ne semble pas être augmenté en réponse directe à une irradiation chronique. Au contraire, il paraît diminuer dans certaines conditions, ce qui pourrait être expliqué par une stimulation importante des systèmes de défense cellulaire, telle la protéolyse, par l’exposition chronique. Cette voie d’investigation devra être prise en compte ainsi qu’une analyse protéomique associée aux conditions expérimentales étudiées pour réaliser un bilan sur ces études. Ceci permettra in fine une amélioration des connaissances sur les marqueurs protéiques possiblement associés à une exposition chronique aux rayonnements ionisants, et en lien avec des effets individuels radioinduits, i.e. reprotoxicité.
Valorisation
Les deux méthodes mises au point pour la mesure des carbonyles ont fait l’objet de publications. D’autres publications sur les résultats sont en cours de rédaction.
Publications :
- Kuzmic M. et al. In situ visualization of carbonylation and its co-localization with proteins, lipids, DNA and RNA in Caenorhabditis elegans, Free Radical Biology and Medicine, 101 (2016) 465–474
- Pyr dit Ruys S. et al. Benchmarking of protein carbonylation analysis in Caenorhabditis elegans: specific considerations and general advice, Free Radical Biology and Medicine, 99(2016)364–373
- Dubois et al. Precoce and opposite response of proteasome activity after acute or chronic exposure of C. elegans to g radiation, Scientific Reports 2018
-
Kuzmic M., Galas S., Lecomte-Pradines C., Dubois C., Dubourg N., Frelon S. Interplay between ionizingradiation effects and aging in C. elegans, Free Radical Biology and Medicine, 2019, Volume 134, 657-665
-
Kuzmic M., Richaud M., Cuq P., Frelon S., Galas S Carbonylation accumulation ofthe Hypsibius exemplaris anhydrobiote reveals age-associated marks, PLoS ONE - Volume 13, Issue 12, December 2018, Article number e0208617
-
Dubois C., Pophillat M., Audebert S., Fourquet P., Lecomte C., Dubourg N., Galas S., Camoin L., Frelon S. Differential modification of the C. elegans proteome in response to acute andchronic gamma radiation: Link with reproduction decline, Science of The Total Environment Volume 676, 1 August 2019, Pages 767-781
Autres :
- Mémoire de thèse C. Dubois ISRN-IRSN-2018-206 - Approche intégrée des dommages des rayonnements ionisants chez Caenorhabditis elegans : de l’ADN aux protéines. Soutenue le 28 novembre 2017 à Cadarache
- Mémoire de thèse M. Kuzmic – Role of protein and DNA damage in biological response to radiation and aging. Soutenue le 12 Juillet 2018 à Split (Croatie)
Dates : 2014-2017
Financements : IRSN, Needs-Environment (projets Memorica et Probiorica sur le « Lien entre la carbonylation des protéines et la reproduction après irradiation gamma aiguë ou chronique de la souche sauvage de C. elegans ».)
Partenaires :
Mediterranean Institute for Life Sciences – Medils (Croatie)